
 Powered by Bioz
Powered by Bioz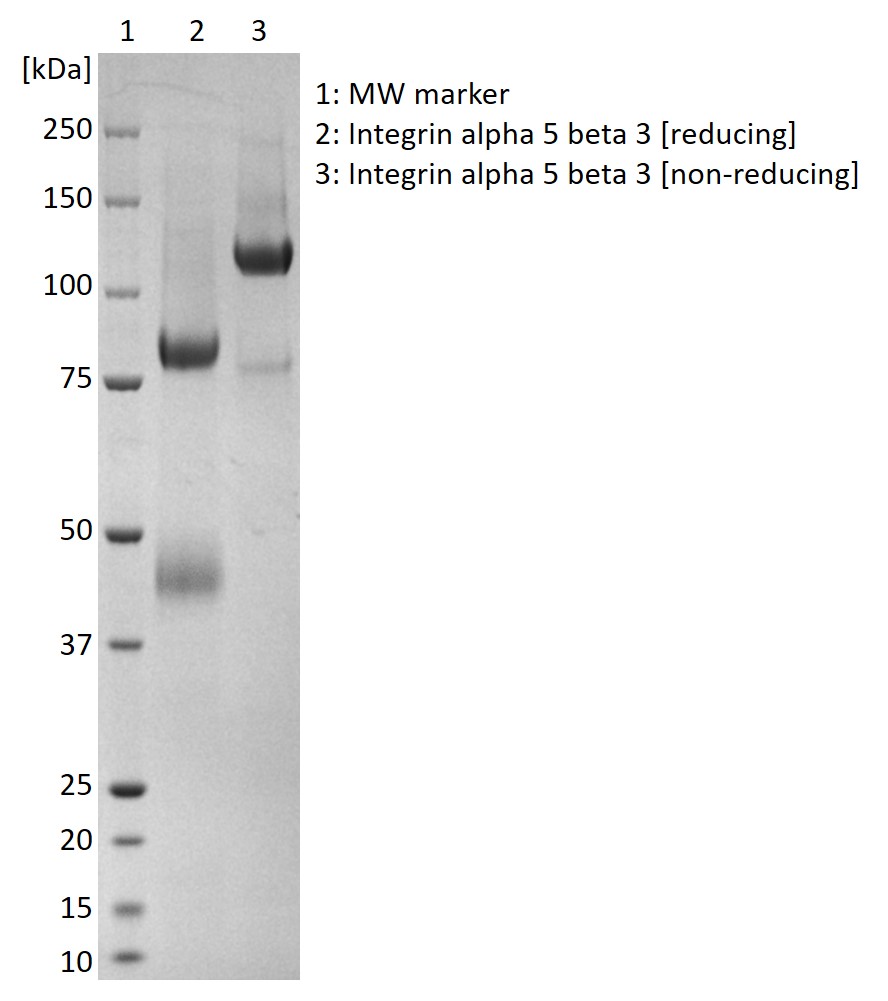
SDS-PAGE: Coomassie-stained SDS-PAGE showing purified alpha-5 beta-3 protein.
Human Integrin Alpha-5 Beta-3, His-Tag
Price range: £492.29 through £1,725.03 excl. VAT
Recombinant human integrin alpha-5 beta-3 protein produced in HEK293 cells and purified from culture supernatant. Protein contains a C-terminal 6x His-tag.
HUMAN INTEGRIN ALPHA-5 BETA-3, HIS-TAG
Recombinant human integrin alpha-5 beta-3 (ITGA5:ITGB3) protein produced in HEK293 cells and purified from culture supernatant.
PRODUCT DETAILS – HUMAN INTEGRIN ALPHA-5 BETA-3, HIS-TAG
- Recombinant human integrin ß-3 protein (NCBI accession number AAA52589.1, AA1-718) and human integrin α-5 (NCBI accession number NP_002196.4, AA1-992).
- Both proteins contain a C-terminal 6x histidine tag.
- Proteins were co-expressed in HEK293 cells and purified from culture supernatant by immobilised metal affinity chromatography.
- Resulting protein is a mix of integrin α-5 and integrin α-5/ß-3 heterodimer, sterile filtered in Dulbecco’s phosphate buffered saline pH7.4.
BACKGROUND
Integrins are heterodimeric integral membrane protein receptors composed of an alpha subunit and a beta subunit that function in cell surface adhesion and signaling. Each subunit crosses the membrane once, with most of the polypeptide residing in the extracellular space, and has two short cytoplasmic domains. A given chain may combine with multiple partners resulting in different integrins. Integrin alpha-5 or CD49e is a protein that in humans is encoded by the ITGA5 gene. Alpha chain 5 undergoes post-translational cleavage in the extracellular domain to yield disulfide-linked light and heavy chains that join with beta 1 to form a fibronectin receptor, recognizing the sequence R-G-D. The integrin alpha 5 and integrin alpha V subunits are encoded by distinct genes. Integrin alpha-V/beta-3 (ITGAV:ITGB3) is a receptor for cytotactin, fibronectin, laminin, matrix metalloproteinase-2, osteopontin, osteomodulin, prothrombin, thrombospondin, vitronectin and von Willebrand factor.
Numerous microorganisms utilize integrins to gain entry into cells. Integrin beta-3 (β3) or CD61 is a protein that in humans is encoded by the ITGB3 gene. Alpha V beta 3 is one of the most promiscuous integrins, as it binds to multiple ligands and also serves as a receptor for several viruses such as foot-and-mouth disease virus, adenovirus, and human immunodeficiency virus (Xiong et al., 2001).
Membrane fusion and attachment of pneumoviruses is achieved by the F protein which contains a putative integrin-binding motif (RGD) and binding to 5β1 and αv integrins are essential for cell-cell fusion and human metapneumovirus (hMPV) infection (Wei et al., 2014).
The specific receptors used for viruses within the Bunyaviridae remain mostly unknown; however, β3 and β1 integrins have been shown to be used as receptors for hantaviruses (Connolly-Andersen et al., 2007). It has been shown that β3 integrins facilitate the cellular entry of pathogenic hantavirus pulmonary syndrome (HPS) associated hantaviruses. Hantaviruses are enveloped viruses and their unique grid-like surface structure may require the use of integrins for entry into cells (Gavrilovskaya et al., 1998).
Other integrins have also been associated with virus entry into cells including ITGA5:ITGB1 which acts as a receptor for human metapneumovirus and as a coreceptor for parvovirus B19 (Weigel-Kelley et al., 2003). Integrin ITGAV:ITGB3 acts as a receptor for Herpes virus 8/HHV-8 (Garrigues et al., 2008), for Coxsackievirus A9 (Roivainen et al., 1994) and Cytomegalovirus/HHV-5 (Wang et al., 2005). Integrin ITGAV:ITGB3 also acts as a receptor for West Nile virus. Integrins were shown to modulate the infection efficiency of West Nile virus into cells and infection was substantially inhibited in Vero cells pretreated with blocking antibodies against αvβ3 integrin and its subunits by receptor competition assay. However, the binding of WNV to αvβ3 integrin does not specifically occur on the RGD binding site. Cells pretreated with antibodies against αvβ3 integrin also inhibited flavivirus Japanese encephalitis and to a lesser extent flavivirus dengue infections (Chu and Ng, 2004; Schmidt et al., 2013).
Antibodies against the receptors αvβ3 and αvβ5 block adenovirus internalization without affecting attachment. Additionally, adenovirus binds to cultured cells lacking αv integrins but fail to become internalized (Wickham et al., 1993).
Integrins may also determine the pathway of virus entry. Herpes Simplex virus (HSV) enters CHO cells lackingαVβ3-integrin through a pathway independent of cholesterol-rich rafts and dynamin2. In the presence of αVβ3-integrin, HSV enters CHO-nectin1 cells through a pathway dependent on cholesterol-rich rafts and dynamin2 (Gianni et al., 2010).
REFERENCES
- Chu and Ng (2004). Interaction of West Nile virus with alpha v beta 3 integrin mediates virus entry into cells. J Biol Chem. 279(52):54533-41.
- Connolly-Andersen et al. (2007). Basolateral entry and release of Crimean-Congo hemorrhagic fever virus in polarized MDCK-1 cells. J Virol. 81(5):2158-64.
- Garrigues et al. (2008). Integrin alphaVbeta3 Binds to the RGD motif of glycoprotein B of Kaposi’s sarcoma-associated herpesvirus and functions as an RGD-dependent entry receptor. J Virol. 82(3):1570-80.
- Gavrilovskaya et al. (1998). beta3 Integrins mediate the cellular entry of hantaviruses that cause respiratory failure. Proc Natl Acad Sci U S A. 95(12):7074-9.
- Gianni et al. (2010). αVβ3-integrin routes herpes simplex virus to an entry pathway dependent on cholesterol-rich lipid rafts and dynamin2. PNAS 107 (51) 22260-22265.
- Horton MA (1997). The alpha v beta 3 integrin vitronectin receptor. Int J Biochem Cell Biol. 29(5):721-5.
- Roivainen et al. (1994). Entry of coxsackievirus A9 into host cells: specific interactions with alpha v beta 3 integrin, the vitronectin receptor. Virology. 1994 Sep;203(2):357-65.
- Schmidt et al. (2013). Integrin ITGAV:ITGB3 acts as a receptor for West nile virus. J Gen Virol. 94(Pt 8):1723-33.
- Wang et al. (2005). Integrin alphavbeta3 is a coreceptor for human cytomegalovirus. Nat Med. 11(5):515-21.
- Wei et al. (2014). Roles of the putative integrin-binding motif of the human metapneumovirus fusion (f) protein in cell-cell fusion, viral infectivity, and pathogenesis. J Virol. 88(8):4338-52.
- Weigel-Kelley et al. (2003). Alpha5beta1 integrin as a cellular coreceptor for human parvovirus B19: requirement of functional activation of beta1 integrin for viral entry. Blood. 102(12):3927-33.
- Wickham et al. (1993). Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell. 73(2):309-19.
- Xiong et al. (2001). Crystal Structure of the Extracellular Segment of Integrin αVβ3. Science. 294(5541): 339–345.