Powered by Bioz
Powered by BiozRABBIT ANTI-BORRELIA BURGDORFERI SENSU STRICTO (B31) DBPB ANTIBODY
This is a polyclonal antibody, prepared against Decorin Binding Protein B (DbpB) from the spirochete B. burgdorferi, for use in ELISA and western blotting applications. Strain B31 is the type strain (ATCC 35210) for this organism and was derived by limited dilutional cloning from the original Lyme-disease tick isolate obtained by A. Barbour (Johnson, et al., 1984). It should be noted that other microbial DbpA’s, such as E. coli (ATP-dependent RNA helicase DbpA), are significantly different to Borrelia DbpA. B. burgdorferi sensu stricto has a linear plasmid (lp54) which carries a two-gene operon encoding two surface lipoproteins, DbpA and DbpB, both of which bind decorin (Guo, et al., 1995).
PRODUCT DETAILS – RABBIT ANTI-BORRELIA BURGDORFERI SENSU STRICTO (B31) DBPB ANTIBODY
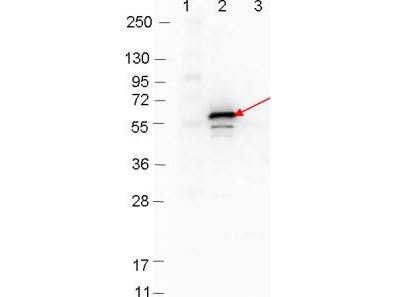
- Rabbit anti-B. burgdorferi sensu stricto DbpB polyclonal IgG antibody (strain B31).
- Greater than 95% purity by SDS-PAGE and buffered in 0.02 M Potassium Phosphate, 0.15 M Sodium Chloride, pH 7.2.
BACKGROUND
DbpA and DbpB are surface-exposed outer membrane lipoproteins that mediate the attachment of Borrelia to decorin, a major component of the host extracellular matrix, enabling bacteria to colonize mammalian tissues (Roberts, et al., 1998). Both can mediate interaction with the glycosaminoglycans (GAGs) heparin and dermatan sulfate (Guo, et al., 1998), but only DbpB binds chondroitin sulfate (Fischer, et al., 2003). The expression of decorin-binding protein (Dbp) A and/or DbpB is sufficient to convert a high-passage nonadherent B. burgdorferi strain into one that efficiently binds 293 epithelial cells.
The spirochete travels from the tick mid-gut during tick feeding, to the tick salivary glands and into the mammal host, and it is believed that this migration is facilitated by changes in expression of different B. burgdorferi genes. It is thought that expression of the various proteins associated with the spirochete may be regulated by the changes in tick life cycle, changes in conditions during tick feeding (such as temperature, pH, and nutrients) and/or in coordination with the course of infection of the mammal host. While not expressed in the unfed tick, DbpA (and possibly DbpB) are quickly upregulated, either during feeding or after entry, into the host. The location of DbpA and DbpB in the outer membrane of B. burgdorferi allows exposure of these proteins to the host immune system. DbpA has also been used to show high levels of genetic diversity in B. afzelii-infected rodent hosts, by qPCR (Coipan, et al., 2018).
NMR and a crystal structure of a DbpA monomer (resolution of 1.60 Å) confirmed three lysines co-localize with decorin to a common basic patch near the C terminus of the protein (Wang & Feng, 2015; Fortune, et al., 2014). Knockout strains confirmed that the lysine residues are required for binding and infection in mice (Fortune, et al., 2014). Strain-specific variations of Borrelia surface proteins also affect tissue tropism (Lin, et al., 2014) as well as differences in GAG binding affinities which is correlated with differences in GAG-binding pocket location and epitope number (Morgan & Wang, 2015).
REFERENCES
- Casselli, T., Tourand, Y. & Bankhead, T., 2012. Altered murine tissue colonization by Borrelia burgdorferi following targeted deletion of linear plasmid 17-carried genes. Infection and Immunity, 80(5), pp. 1773-1782.
- Coipan, C. E. et al., 2018. The genetic diversity of Borrelia afzelii is not maintained by the diversity of the rodent hosts. Parasit Vectors, 11(1), p. 454.
- Fischer, J. R., Parveen, N., Magoun, L. & Leong, J. M., 2003. Decorin-binding proteins A and B confer distinct mammalian cell type-specific attachment by Borrelia burgdorferi, the Lyme disease spirochete. Proc. Natl. Acad. Sci. U. S. A. , Volume 100, p. 7307–7312.
- Fortune, D. E. et al., 2014. Identification of Lysine Residues in the Borrelia burgdorferi DbpA Adhesin Required for Murine Infection. Infect Immun., 82(8), p. 3186–3198.
- Guo, B. P. et al., 1998. Decorin-binding adhesins from Borrelia burgdorferi. Mol. Microbiol. , Volume 30, p. 711–723.
- Guo, B. P., Norris SJ, S. J., Rosenberg, L. C. & Hook, M., 1995. Adherence of Borrelia burgdorferi to the proteoglycan decorin. Infect. Immun., Volume 63, p. 3467–3472.
- Johnson, R.C., et al. 1984. Borrelia burgdorferi sp. nov.: etiologic agent of Lyme disease. Int J Syst Bacteriol, 34, pp. 496–497.
- Lin, Y. P., Benoit, V., Yang, X. & Martínez-Herranz, R., 2014. Strain-specific variation of the decorin-binding adhesin DbpA influences the tissue tropism of the lyme disease spirochete. PLoS Pathog. , 10(7), p. 14.
- Morgan, A. M. & Wang, X., 2015. Structural mechanisms underlying sequence-dependent variations in GAG affinities of decorin binding protein A, a Borrelia burgdorferi adhesin. Biochem J., 467(3), pp. 439-51.
- Roberts, W. C., Mullikin, B. A., Lathigra, R. & Hanson, M. S., 1998. Molecular analysis of sequence heterogeneity among genes encoding decorin binding proteins A and B of Borrelia burgdorferi sensu lato. Infect. Immun., Volume 66, p. 5275–5285.
- Wang, X. & Feng, W., 2015. Structure of decorin binding protein B from Borrelia burgdorferi and its interactions with glycosaminoglycans. Biochim Biophys Acta., 1854(12), pp. 1823-1832.