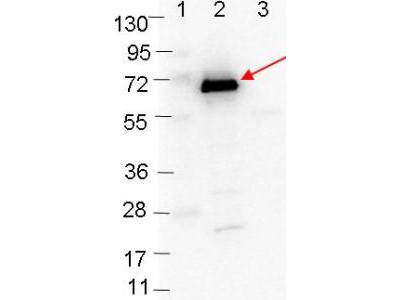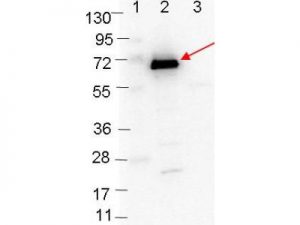
Western blot showing detection of 0.1µg of recombinant OspC protein. Lane 1: Molecular weight markers. Lane 2: MBP-OspC fusion protein (arrow; expected MW: 63.1 kDa). Lane 3: MBP alone. Protein was run on a 4-20% gel, then transferred to 0.45 µm nitrocellulose. After blocking with 1% BSA-TTBS overnight at 4°C, primary antibody was used at 1:1000 at room temperature for 30 min. HRP-conjugated Goat-Anti-Rabbit secondary antibody was used at 1:40,000 in HiGlo Blocking Buffer and imaged on the VersaDoc™ MP 4000 imaging system (Bio-Rad).
RABBIT ANTI-BORRELIA BURGDORFERI SENSU STRICTO (B31) OSPC ANTIBODY
Rabbit Anti-Borrelia burgdorferi OspC antibody, is a polyclonal suitable for use in ELISA and Western blotting applications.
PRODUCT DETAILS – RABBIT ANTI-BORRELIA BURGDORFERI SENSU STRICTO (B31) OSPC ANTIBODY
- Rabbit anti-B. burgdorferi sensu stricto OspC polyclonal IgG antibody (strain B31).
- Greater than 95% purity by SDS-PAGE and buffered in 0.02 M Potassium Phosphate, 0.15 M Sodium Chloride, pH 7.2.
BACKGROUND
Strain B31 is the type strain (ATCC 35210) for this organism and was derived by limited dilutional cloning from the original Lyme-disease tick isolate obtained by A. Barbour (Johnson, et al., 1984).
Outer Surface Protein C, or OspC, is a 22 kDa immunogenic protein on the outer surface of the spirochete, B. burgdorferi (Caine, et al., 2017). OspC’s function is not known, but it is located by lipid-anchoring sites on the outer cell membrane, although it is not believed to be associated with lipid rafts (Grimm, et al., 2004; Toledo, et al., 2014).
OspC has been shown to be a requisite for B. burgdorferi to establish mammalian infection as well as for spirochaetal transmission from ticks to mammals. Expression begins during blood feeding by the nymphal tick and continues to be produced in the early stages of infection in the mammal host and is highly immunogenic in mice. As one of the strategies to evade host humoral responses, spirochetes downregulate OspC production in response to anti-OspC antibodies within 2 to 3 weeks after infection in mice (Grimm, et al., 2004).
OspC also has been proposed to play roles in promoting survival and/or dissemination of spirochetes within the mammalian host. For example, OspC binds to a tick salivary protein, Salp15, which can protect spirochetes from complement- and antibody-mediated killing (Hovius, et al., 2008). OspC also binds host plasminogen and this phenotype correlates with invasiveness of spirochetes in mice (Tilly, et al., 2006) and has antiphagocytic properties, aiding spirochete evasion of mononuclear phagocytes during early infection in mammals (Carrasco, et al., 2015).
OspC is required for B. burgdorferi infection in both immunocompetent and SCID mice (lacking B and T cells) and has been proposed to facilitate evasion of innate immune defences. An OspC knockout mutant was cleared within the first 48 h of infection in a murine host, suggesting a protective role of OspC against innate defences. Constitutive expression of heterologous lipoproteins in the OspC mutant was shown to restore infection in SCID mice, suggesting that OspC may also play a nonspecific structural role for B. burgdorferi (Smith, et al., 2018).
REFERENCES
- Caine, J. A. et al., 2017. Borrelia burgdorferi outer surface protein C (OspC) binds complement component C4b and confers bloodstream survival. Cell Microbiol., Volume 12, p. 19.
- Carrasco, S. E. et al., 2015. Outer Surface Protein OspC Is an Antiphagocytic Factor That Protects Borrelia burgdorferi from Phagocytosis by Macrophages. Infection and Immunity, Volume 83.
- Grimm, D. et al., 2004. Outer-surface protein C of the Lyme disease spirochete: a protein induced in ticks for infection of mammals. Proc. Natl. Acad. Sci. U. S. A., Volume 10.
- Hovius , J. W. et al., 2008. Preferential protection of Borrelia burgdorferi sensu stricto by a Salp15 homologue in Ixodes ricinus saliva. J Infect Dis., 198(8), pp. 1189-97.
- Johnson, R.C., et al. 1984. Borrelia burgdorferi sp. nov.: etiologic agent of Lyme disease. Int J Syst Bacteriol, 34, pp. 496–497.
- Smith, T. C. et al., 2018. Borrelia host adaptation Protein (BadP) is required for the colonization of a mammalian host by the agent of Lyme disease. Infect Immun. .
- Tilly, K. et al., 2006. Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect. Immun. , Volume 74, p. 3554–3564.
- Toledo, A. et al., 2014. Selective Association of Outer Surface Lipoproteins with the Lipid Rafts of Borrelia burgdorferi. mBio, 5(2).


 Powered by Bioz
Powered by Bioz